Post by brobear on Mar 31, 2020 3:40:04 GMT -5
esajournals.onlinelibrary.wiley.com/doi/full/10.1890/06-0624.1
Arctic Marine Mammals
Polar bear (Ursus maritimus)
Polar bears are similar to brown bears (Ursus arctos) in size and mass but are generally longer and slenderer. They have a downturned rather than dished face. Adult males weigh 350–650 kg, and adult females weigh approximately 150–300 kg. They are covered with whitish fur except for the nose and footpads. Canine teeth are well developed, and cheek teeth are more trenchant than those of other bears, reflecting evolution from an omnivorous to carnivorous diet (Kurtén 1964).
Arctic adaptations include: white coats that help camouflage them and aid in stalking seals; dense fur and thick body fat, providing insulation from cold air and water (further, body fat helps buoy up the animal); a streamlined teardrop body shape and large, oar‐like paws that enable them to swim well and the paws act like snowshoes on thin, newly formed ice; short furry ears and large body size that help to conserve heat; an extremely well‐developed sense of smell that aids in finding food and bringing together the sexes over vast icy expanse during the breeding season; the ability to make dens in coastal snowbanks, providing protection for the mother and her (usually) twin cubs during the coldest period of the year; hunting techniques such as sniffing out and breaking into seal dens and waiting by seal breathing holes.
Polar bear fossils are confined to the Pleistocene (Kurtén 1964). Both fossil and molecular data indicate that polar bears stemmed from brown bears about the Middle Pleistocene (perhaps 400000–300000 years ago) (Kurtén 1964, Talbot and Shields 1996). Middle Pleistocene climatic cooling probably influenced the evolution of the ancestor of the polar bear (Kahlke 1999). Yu et al. (2004), by combining nuclear and mitochondrial DNA findings, have gained new insight into the evolutionary history of the bears (Ursidae). Their results corroborate other morphologic and genetic evidence indicating that brown and polar bears are most closely related and suggest that polar bears split from brown bears between 1 and 1.5 million years ago (Ma).
The divergence of polar and brown bears probably occurred somewhere along the coasts of Siberia or Alaska, but pre‐Holocene fossils are scarce. Heaton et al. (1996) note that differences in mitochondrial DNA sequences suggest that living brown bears of the Alexander Archipelago, southeastern Alaska, comprise a distinct clade and are most closely related to polar bears. Could that be a clue as to their place of origin? I speculate that the drift from an Arctic coastal brown bear with a grayish coat to a polar bear began with the former specializing in scavenging beached marine mammal carcasses. Gradually, the bears might have ranged farther out onto the sea ice (fast ice) where they could actively prey on young seals in their dens and then on adult ringed seals at their breathing holes. In this respect, it is worth noting that grizzly bears (Ursus arctos) are now regular visitors to the Canadian Arctic Islands. On the sea ice of Viscount Melville Sound, remains of seals as well as female polar bears and their cubs have been attributed to kills by grizzly bears. Perhaps one of these bears denned on Melville Island during the winter of 2003–2004 (Doupé et al. 2007). Breakthroughs would come with genetic selection for white coats (camouflage) to aid their hunting (Sterling 1989:26), as well as adaptation to denning in dense snow near sea coasts rather than in earth farther inland, and physical adaptation for more trenchant teeth to support the more carnivorous diet, as well as large, oar‐like feet to enhance their swimming ability.
The oldest known polar bear fossils are a relatively large right ulna from presumably Early Weichselian (approximately 70000 BP[?]) gravels of the Thames at Kew Bridge, London, and a left lower jaw from >80000 BP deposits on Svalbard (O. Ingólfsson, personal communication).2 The Kew Bridge find induced Kurtén (1964) to create a new gigantic Late Pleistocene subspecies Ursus maritimus tyrannus. Stuart (1982) calculated that the Kew locality was at least 140 km from the Devensian (last glaciation) coastline, based on the assumption that sea level was at its lowest then. Further, the presence of a marine mammal fauna, including ringed and bearded seals, suggests that polar bears too were present in the southern North Sea during the Late Pleistocene (at several colder intervals during the Weichselian) (Post 2005). Also, it is possible that outlines of bears in the Paleolithic cave of Ekain, northern Spain, may indicate that polar bears drifted south from the edge of pack ice off southern England (see map on the cover of Preece [1995]) to the northern coast of Spain during heavy ice years toward the close of the Pleistocene. I interpret the bears to be twin subadult polar bears from their teardrop body shapes (Bahn and Vertut 1988: Fig. 90). These images were probably made during the Magdalenian (approximately 17000–12000 years ago).
The latest Weichselian to Early Holocene colonization of the western coastal regions of Norway, Sweden, and Denmark (northern Jutland) are well documented. A large lower jaw of an adult male polar bear dated to 11100 ± 160 BP from near Asdal (Jutland) occurs in the Allerød (Aaris‐Sørensen and Petersen 1984). Eight finds (from Scania [two], Halland [one], and Bohuslän [five]) are known from Sweden (Kurtén 1964, Liljegren 1975). The more southerly specimens from Scania are dated at 12710 BP and 12490 BP (Håkansson 1974, 1976). Two of the finds from Bohuslän have been dated to 10620 BP and 10430 BP (Håkansson 1976). A nearly complete skeleton of a large male nearly 30 years old from Judaberg (Finnøy, Norway) has been dated at 10660 BP. It shows that polar bears were present in the Younger Dryas High Arctic marine environment in southwestern Norway (Blystad et al. 1983). Otto Salvigsen (personal communication) has dated (about 8200 BP) polar bear bones from a marine section on Svalbard. Fossil finds from Siberia are rare and not well dated. Vereshchagin (1969) records a right ulna from near the mouth of Mordy‐Yahk River on the western coast of the Yamal Peninsula. Fossil remains of walrus and white whale were found there too. Also identified as belonging to the polar bear are: a right first molar stained black (Pleistocene?) found in 1961 on the shore of the Pechora River, as well as abundant remains from the Mesolithic site on Zhokhov Island (approximately 9000–8000 BP) (G. F. Baryshnikov, personal communication). The early inhabitants of Zhokhov Island in the Siberian Arctic evidently relied heavily upon polar bears for subsistence. Many of the bones from the site exhibit cut marks. Skull measurements suggest that female bears with cubs were killed in winter by hunters seeking their dens (Pitulko and Kasparov 1996, Pitulko 2003). In northern North America, the oldest radiocarbon‐dated specimens from Vandfeldsnaes, Greenland, and Cape Richard Collinson, Prince of Wales Island, Canada, are only about 2000 years old (Harington 2003a:383).
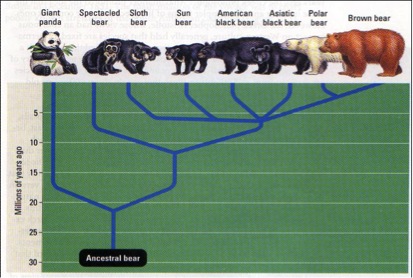
Fulton and Strobeck (2006: Fig. 3a) provide a succinct view of relationships among bears. Note particularly the closeness and relatively recent evolution of polar and brown bears (Fig. 2).
Relationships among the bears according to a maximum‐likelihood cladogram recovered from all genes (Fulton and Strobeck 2006: Fig. 3a). Note the relatively recent divergence of polar and brown bears. Values above the branches are maximum parsimony (MP) bootstrap/Bayesian posterior probability (BPP) values. The nodes that do not have support values were polytomous in both topologies.
Modern and fossil distributions of polar bears are given in Vereshchagin (1969:46, Fig. 6) and Laidre et al. (2008).

Arctic Marine Mammals
Polar bear (Ursus maritimus)
Polar bears are similar to brown bears (Ursus arctos) in size and mass but are generally longer and slenderer. They have a downturned rather than dished face. Adult males weigh 350–650 kg, and adult females weigh approximately 150–300 kg. They are covered with whitish fur except for the nose and footpads. Canine teeth are well developed, and cheek teeth are more trenchant than those of other bears, reflecting evolution from an omnivorous to carnivorous diet (Kurtén 1964).
Arctic adaptations include: white coats that help camouflage them and aid in stalking seals; dense fur and thick body fat, providing insulation from cold air and water (further, body fat helps buoy up the animal); a streamlined teardrop body shape and large, oar‐like paws that enable them to swim well and the paws act like snowshoes on thin, newly formed ice; short furry ears and large body size that help to conserve heat; an extremely well‐developed sense of smell that aids in finding food and bringing together the sexes over vast icy expanse during the breeding season; the ability to make dens in coastal snowbanks, providing protection for the mother and her (usually) twin cubs during the coldest period of the year; hunting techniques such as sniffing out and breaking into seal dens and waiting by seal breathing holes.
Polar bear fossils are confined to the Pleistocene (Kurtén 1964). Both fossil and molecular data indicate that polar bears stemmed from brown bears about the Middle Pleistocene (perhaps 400000–300000 years ago) (Kurtén 1964, Talbot and Shields 1996). Middle Pleistocene climatic cooling probably influenced the evolution of the ancestor of the polar bear (Kahlke 1999). Yu et al. (2004), by combining nuclear and mitochondrial DNA findings, have gained new insight into the evolutionary history of the bears (Ursidae). Their results corroborate other morphologic and genetic evidence indicating that brown and polar bears are most closely related and suggest that polar bears split from brown bears between 1 and 1.5 million years ago (Ma).
The divergence of polar and brown bears probably occurred somewhere along the coasts of Siberia or Alaska, but pre‐Holocene fossils are scarce. Heaton et al. (1996) note that differences in mitochondrial DNA sequences suggest that living brown bears of the Alexander Archipelago, southeastern Alaska, comprise a distinct clade and are most closely related to polar bears. Could that be a clue as to their place of origin? I speculate that the drift from an Arctic coastal brown bear with a grayish coat to a polar bear began with the former specializing in scavenging beached marine mammal carcasses. Gradually, the bears might have ranged farther out onto the sea ice (fast ice) where they could actively prey on young seals in their dens and then on adult ringed seals at their breathing holes. In this respect, it is worth noting that grizzly bears (Ursus arctos) are now regular visitors to the Canadian Arctic Islands. On the sea ice of Viscount Melville Sound, remains of seals as well as female polar bears and their cubs have been attributed to kills by grizzly bears. Perhaps one of these bears denned on Melville Island during the winter of 2003–2004 (Doupé et al. 2007). Breakthroughs would come with genetic selection for white coats (camouflage) to aid their hunting (Sterling 1989:26), as well as adaptation to denning in dense snow near sea coasts rather than in earth farther inland, and physical adaptation for more trenchant teeth to support the more carnivorous diet, as well as large, oar‐like feet to enhance their swimming ability.
The oldest known polar bear fossils are a relatively large right ulna from presumably Early Weichselian (approximately 70000 BP[?]) gravels of the Thames at Kew Bridge, London, and a left lower jaw from >80000 BP deposits on Svalbard (O. Ingólfsson, personal communication).2 The Kew Bridge find induced Kurtén (1964) to create a new gigantic Late Pleistocene subspecies Ursus maritimus tyrannus. Stuart (1982) calculated that the Kew locality was at least 140 km from the Devensian (last glaciation) coastline, based on the assumption that sea level was at its lowest then. Further, the presence of a marine mammal fauna, including ringed and bearded seals, suggests that polar bears too were present in the southern North Sea during the Late Pleistocene (at several colder intervals during the Weichselian) (Post 2005). Also, it is possible that outlines of bears in the Paleolithic cave of Ekain, northern Spain, may indicate that polar bears drifted south from the edge of pack ice off southern England (see map on the cover of Preece [1995]) to the northern coast of Spain during heavy ice years toward the close of the Pleistocene. I interpret the bears to be twin subadult polar bears from their teardrop body shapes (Bahn and Vertut 1988: Fig. 90). These images were probably made during the Magdalenian (approximately 17000–12000 years ago).
The latest Weichselian to Early Holocene colonization of the western coastal regions of Norway, Sweden, and Denmark (northern Jutland) are well documented. A large lower jaw of an adult male polar bear dated to 11100 ± 160 BP from near Asdal (Jutland) occurs in the Allerød (Aaris‐Sørensen and Petersen 1984). Eight finds (from Scania [two], Halland [one], and Bohuslän [five]) are known from Sweden (Kurtén 1964, Liljegren 1975). The more southerly specimens from Scania are dated at 12710 BP and 12490 BP (Håkansson 1974, 1976). Two of the finds from Bohuslän have been dated to 10620 BP and 10430 BP (Håkansson 1976). A nearly complete skeleton of a large male nearly 30 years old from Judaberg (Finnøy, Norway) has been dated at 10660 BP. It shows that polar bears were present in the Younger Dryas High Arctic marine environment in southwestern Norway (Blystad et al. 1983). Otto Salvigsen (personal communication) has dated (about 8200 BP) polar bear bones from a marine section on Svalbard. Fossil finds from Siberia are rare and not well dated. Vereshchagin (1969) records a right ulna from near the mouth of Mordy‐Yahk River on the western coast of the Yamal Peninsula. Fossil remains of walrus and white whale were found there too. Also identified as belonging to the polar bear are: a right first molar stained black (Pleistocene?) found in 1961 on the shore of the Pechora River, as well as abundant remains from the Mesolithic site on Zhokhov Island (approximately 9000–8000 BP) (G. F. Baryshnikov, personal communication). The early inhabitants of Zhokhov Island in the Siberian Arctic evidently relied heavily upon polar bears for subsistence. Many of the bones from the site exhibit cut marks. Skull measurements suggest that female bears with cubs were killed in winter by hunters seeking their dens (Pitulko and Kasparov 1996, Pitulko 2003). In northern North America, the oldest radiocarbon‐dated specimens from Vandfeldsnaes, Greenland, and Cape Richard Collinson, Prince of Wales Island, Canada, are only about 2000 years old (Harington 2003a:383).
Fulton and Strobeck (2006: Fig. 3a) provide a succinct view of relationships among bears. Note particularly the closeness and relatively recent evolution of polar and brown bears (Fig. 2).
Relationships among the bears according to a maximum‐likelihood cladogram recovered from all genes (Fulton and Strobeck 2006: Fig. 3a). Note the relatively recent divergence of polar and brown bears. Values above the branches are maximum parsimony (MP) bootstrap/Bayesian posterior probability (BPP) values. The nodes that do not have support values were polytomous in both topologies.
Modern and fossil distributions of polar bears are given in Vereshchagin (1969:46, Fig. 6) and Laidre et al. (2008).